Hereditary Deafness in Dogs and Cats: Causes, Prevalence, and Current Research
George M. Strain
Comparative Biomedical Sciences, School of Veterinary Medicine, Louisiana State University
Baton Rouge, LA, USA
Hearing loss is the most common sensory deficit in humans, and may also be so in companion animals where it affects all mammalian species. Deafness in an animal is usually not a life-threatening disorder in the way of some renal disorders, and it is not a painful condition like hip dysplasia, but it does put an animal at risk from undetected dangers such as motor vehicles or predators, and deaf animals create their own liabilities and present non-trivial training challenges to their owners. Because large numbers of deaf puppies and kittens get put down, it is important to identify animals affected by the hereditary forms of deafness B both unilaterally and bilaterally deaf B and remove them from the potential breeding pool to reduce the number of future deaf animals.
Anatomy and Physiology
The ear has three compartments: (1) the outer ear, including the pina and the ear canal down to the tympanic membrane, (2) the air-filled middle ear, including the three ossicles (malleus, incus, and stapes) and the connection to the pharynx (the auditory canal), and (3) the fluid-filled inner ear or cochlea, which contains the organ of Corti and the initial portion of the auditory nerve. The cochlea is imbedded in solid bone, which protects it from all but the most severe trauma. Air vibrations (sound) in the ear canal vibrate the tympanum; these oscillations are amplified and transmitted through the three ossicles to a membranous opening (the oval window) into the cochlea. Oscillations of the oval window are transmitted through the fluids of the snail-shaped cochlea, causing deflections of small stereocilia on the sensory hair cells of the organ of Corti, thereby initiating nerve impulses into the brain
Click on image to see a larger view
| Figure 1. | 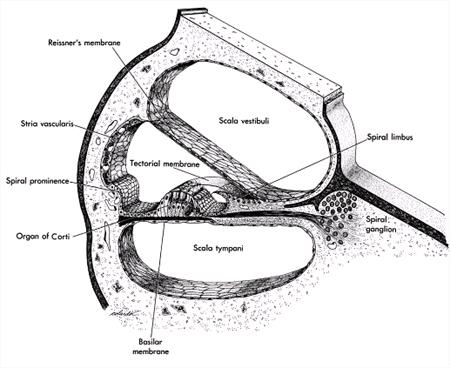
|
|
| |
The organ of Corti wraps around in the cochlea in a fluid-filled compartment. That fluid is high in potassium and low in sodium, similar to intracellular fluid. On the outer wall of this compartment is a special vascular bed, the stria vascularis, that is responsible for maintaining the high K+ and low Na+ levels. Part of the mechanism by which the stria for maintains these ionic levels involves pigment cells B melanocytes.1 This structure is a major target of mechanisms producing deafness.
Forms of Deafness
Forms of deafness can be sorted into three complimentary categories, each with two types. Deafness can be:
 inherited or acquired,
inherited or acquired,
congenital or later-onset, or
sensorineural or conductive.
This results in eight possible combinations, such as acquired later-onset sensorineural deafness. Sensorineural (nerve) deafness is loss of auditory function because of loss of cochlear hair cells or auditory nerve neurons. Conductive deafness is blockage of sound transmission through the outer and/or middle ear without damage to the cochlea. Most congenital deafness is hereditary, and most later-onset deafness (as far as we know) is acquired, although there are human forms of inherited later-onset deafness. The most commonly seen forms of deafness in companion animals are (1) hereditary congenital sensorineural deafness, (2) acquired later-onset sensorineural deafness, and (3) acquired later-onset conductive deafness. Acquired conductive deafness may result from otitis externa, otitis media, excess cerumen production, or congenital defects in the ossicles (rare). Acquired sensorineural deafness may result from otitis interna, meningitis, drug toxicity (e.g., gentamicin), noise trauma, mechanical trauma, anoxia, anesthesia, or aging (presbycusis). In human audiology deafness is also classified as syndromic (associated with other disorders) or nonsyndromic. Allport syndrome, which presents with both sensorineural deafness and kidney disease, is an example.
Finally, sensorineural deafness can be primary or secondary. Primary deafness results from destruction of hair cells in the cochlea without antecedent events. This occurs in hereditary deafness in Doberman pinchers, some forms of ototoxicity, and presbycusis. Secondary deafness occurs when hair cells die as a consequence of other damage in the cochlea, most commonly to the stria vascularis. This occurs in pigment-associated hereditary deafness and some forms of ototoxicity. Degeneration of the stria vascularis eliminates the high K+ levels in the fluids surrounding the hair cells, resulting in death of the hair cells and deafness. In pigment-associated hereditary deafness, the strial degeneration and hair cell death usually occur 2-4 weeks after birth.
Hearing Testing
Behavioral testing of hearing is usually accomplished by making a sound outside the animal's visual field, or while the animal is sleeping, and observing for any response. Lack of a response may be interpreted as bilateral deafness, but it may instead reflect an overly stressed animal, an inattentive one, or one who has tired of this game. A response may indicate that at least one ear hears, but deaf animals are especially attentive to other sensory cues, so it may respond from detection of visual, vibration, or air current stimuli. A unilaterally deaf animal responds to sound stimuli B the only behavioral deficit is a difficulty localizing the location of a sound source, so it may turn in the wrong direction to a stimulus. As a result, behavioral testing is subjective and limited in its reliability.
Electrodiagnostic testing provides an objective, non-invasive assessment of the presence or absence of auditory function, and is the gold standard of auditory testing in animals. It permits detection of unilateral deafness and can be performed in awake or anesthetized animals, but facilities for this testing may not be locally available. The hearing test is known as thebrainstem auditory evoked response (BAER) [also known as the brainstem auditory evoked potential (BAEP) and auditory brainstem response (ABR)]. It detects electrical activity in the cochlea and auditory pathways in the brain in much the same way that an EKG detects the electrical activity of the heart.2-4 The response waveform consists of a series of peaks identified with Roman numerals (Figure 2, top): peak I is produced by the cochlea and auditory nerve, and later peaks are produced within the lower brain (brainstem).
Click on image to see a larger view
| Figure 2. | 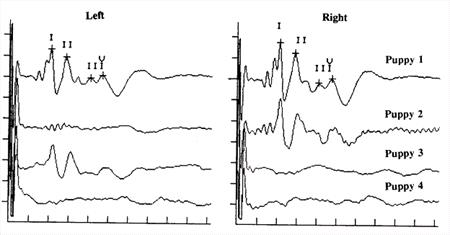
|
|
| |
The response from an ear that is deaf is an essentially flat line. In the sample recordings shown in Figure 2, Puppy 1 heard in both ears, Puppies 2 and 3 were deaf in one ear, and Puppy 4 was deaf in both ears. Because the response amplitude is very small it is necessary to average the responses to multiple stimuli (clicks) to unmask them from the other unrelated electrical activity that is also present on the scalp (EEG, muscle activity, etc).
The response is collected with a special computer through small needle electrodes placed under the skin of the scalp: one in front of each ear, one at the top of the head, and a ground. It is rare for a dog to show any evidence of pain from the placement of the electrodes B if anything the dog objects to the gentle restraint and the irritation of wires hanging in front of its face. The stimulus click produced by the computer is directed into the ear with a foam insert earphone. Each ear is tested individually, and the test usually is complete in 10-15 minutes. Sedation or anesthesia are not necessary unless the dog becomes extremely agitated, which can usually be avoided with patient and gentle handling. A printout of the test results, showing the actual recorded waveform, is provided at the end of the procedure. Because of the time after birth at which congenital deafness develops (2-4 weeks), testing can be performed any time starting at 5 weeks of age. Since there is no known hereditary late-onset deafness in dogs or cats, a single normal recording is good for the animal=s life B barring error, any later hearing loss is assumed to be acquired and not hereditary. BAER tests do not provide a quantitative assessment of hearing loss (i.e., 40 dB) or frequency-specific results, but instead primarily are used to identify totally deaf ears. The click stimulus simultaneously activates receptors for most of the frequencies detected by dogs and cats; the click is similar to white light, which contains all colors. A list of BAER test sites is maintained at www.lsu.edu/deafness/baersite.htm.5
When there is reason to suspect conductive deafness (ongoing or chronic otitis) and the BAER response is flat, the test can be repeated using a vibratory stimulus transducer known as a bone stimulator instead of the air-conducted click produced by the insert earphones. The same stimulus signal is sent to the bone stimulator as to the earphones by the electrodiagnostic instrument, producing a brief vibration. The bone stimulator is firmly held against the bone of the skull at the mastoid process behind the ear. The stimuli are conducted directly through the bone of the skull to the cochlea (which is embedded in the mastoid bone), bypassing the outer and middle ears. If the cochlea is functional, a response will be detected that is very similar to that produced in a normally hearing animal in response to air-conducted stimuli. This test is more difficult to perform because of the difficulty of establishing and maintaining a good mechanical contact between the transducer and bone, especially in heavily muscled dogs. Some dogs show discomfort from the necessary intense pressure against the head, so air-conducted testing of both ears is completed before performing bone testing. Interpretation of the results from bone-conducted stimuli is more subjective than with air-conducted stimuli because of the greater difficulty in obtaining satisfactory recordings. Unfortunately, not all BAER test sites are equipped to perform bone stimulation BAER tests. Conduction deafness is rare in very young puppies, but a normal BAER from bone stimulator testing can keep a hearing dog from being falsely diagnosed with deafness and thereby eliminated from breeding consideration.
Pigment Genes and Hereditary Congenital Sensorineural Deafness
An association between deafness and blue-eyed white cats was noted as early as 1828, and Darwin commented on it in his famous publication The Origin of Species in 1859. Blue-eyed Dalmatians were noted for having deafness as early as 1896. So, the existence of a relationship between white pigmentation and deafness in dogs and cats is not new, and there is an extensive bibliography on the subject,6-12 but the mechanism behind the relationship has only recently begun to be understood.
Melanocytes, which produce pigment granules in skin, hair, and elsewhere, originate embryologically in the neural crest, the source of all neural cells, explaining the linkage between pigment and a neurologic disorder. Melanocytes produce pigment granules B either eumelanin (black or brown) or pheomelanin (yellow or red) B from the amino acid tyrosine. Albinism, in which melanocytes are present but one of the enzymes responsible for melanin production (tyrosinase) is absent or diminished, does not have an association with deafness. Otherwise, white color results from an absence of melanin, usually from an absence of melanocytes. Specific genes have been identified that produce white by suppressing melanocytes B either their migration from the neural crest, their maturation upon arrival at their final site, or their survival. Other genes are responsible for the pigment colors in the pigmented areas. In dogs the genes for white are the recessive alleles of the piebald gene (S) or the dominant allele of the merle gene (M).13,14 In cats the genes are the dominant allele of the white gene (W) and possibly the recessive piebald gene (S).13 In turn, the effects of these genes can be modified by other, as yet unidentified, genes that can result in strong gene expression or weak gene expression (see below).
The S locus in dogs has at least four alleles. The dominant allele S is known as self or non-spotted, and produces a completely pigmented body surface, although minor areas of white may be present on the feet or thorax.
The si allele produces Irish spotting and presents with only a few white areas that are usually on locations such as the thorax, feet, face, or head.
The sp allele produces piebald spotting and produces significantly more white on the body surface than Irish spotting, including the limbs.
The sw or extreme-white piebald allele is associated with an even greater extent of white pigmentation, including the ears and base of the tail.
These alleles are listed in order of decreasing dominance. Although the specific allele responsible for white pigmentation is not known for many breeds, the Basenji and bloodhound are examples of homozygous Irish spotting, the beagle is an example that is usually homozygous piebald, and the Dalmatian and white bull terrier are examples of homozygous extreme-white piebald. Because these alleles are recessive they must be present in pairs to produce their white pattern, but it is possible for a dog to carry one copy each of two of the recessive alleles, as may occur with Boston terriers that are normally sisi but that may on occasion be sisp or sisw.13
The alleles sp and sw are present in the great majority of breeds recognized to be subject to congenital deafness when the identity of pigmentation genes is known, but the white-producing allele is often not known. It is likely that additional genes regulate the expression of the three recessive S alleles. For example, strong expression of sw in Dalmatians results in blue irises from suppression of melanocytes in the eye, and weak expression of sw results in the large pigmented area on some Dalmatians known as a patch, which is present at birth when the rest of the puppy is still white.
A second canine pigmentation locus associated with deafness is merle (M). Homozygosity of the recessive allele (mm) produces uniform pigmentation, while the heterozygous merle (Mm) produces dappling or alternate body areas of fully pigmented coat and pale eumelanic or even white coat. Homozygous merles (MM) are usually nearly solid white, and in some breeds may be deaf, blind with microphthalmic eyes, and sterile. Dogs heterozygous for M are variable in their likelihood of deafness. The harlequin gene (HH) has been identified as a dominant modifier of the merle gene in Great Danes that is lethal when present in the homozygous state;15 harlequin Danes are at elevated risk for deafness, while other color variants are less likely to be affected. Great Dane dogs may carry M, m, HH, si, sp, or sw gene alleles in various combinations, as well as other potential modifier genes, which provides an indication of the complexity of pigmentation genetics in various dog breeds.
Other genes reported to produce white or light coat color in dogs B flecking, ticking, dilution with fawn B do not appear to be associated with deafness.
The W gene has been studied exclusively in domestic longhair and shorthair cats that were not from identifiable or identified breeds; there do not appear to have been deafness studies in any cat pure breed. The white pigment gene in cats is autosomal dominant over color (w), and is unrelated to albinism. Cats with this gene are not always solid white, often having a colored spot on the head that fades with age. Unlike homozygous merle dogs, homozygous W cats do not have clear-cut visual or reproductive deficits, although white cats had a higher mortality than colored cats (32% vs. 26%) during a 30-day quarantine period in one study16 and embryo viability is lower in white than non-white cats.17 Homozygous white cats are more prone to blue eyes (and deafness), and the likelihood of deafness increases with the number of blue eyes.6 White cats carrying the underlying cs Siamese dilution pigment gene can have blue eyes without deafness, which is suggested as one reason why pure-breed white cats may be deaf less often than mixed-breed white cats;18 however, since there are no studies of the prevalence of deafness in pure-breed cats this assertion about differences in prevalence cannot be validated. Long-haired white cats were reported to have a higher prevalence of blue eyes and deafness than short-haired cats,7 but it is not certain whether this observation would stand up in a larger study.
A piebald gene (S) is also said to be found in various cat breeds17,18 that produces white spotting, especially on the neck, feet, and belly, but its existence is not established. There is no report of deafness associated with its presence.
In dogs and cats with white-producing genes, deafness appears to result from strong expression of the gene. As stated above, melanocytes in the stria vascularis play a role in maintaining the ionic environment needed by the cochlear hair cells. When the piebald, merle, or white gene is strongly expressed, it suppresses melanocytes not only in the skin, but also in the iris (a blue eye is missing pigment granules, and the blue results for the same reason the sky is blue) and the stria. When the strial melanocytes are absent the stria degenerates, after which the hair cells die and the various cochlear structures collapse and the auditory nerve fibers start to degenerate. Not all blue-eyed animals are deaf, and all deaf animals don't have blue eyes, but there is a strong statistical correlation between the two.11,12 Conversely, weak expression of the gene is associated with a reduced likelihood of deafness. Dalmatians have a base color of black (dominant) or liver (recessive) covered up by the recessive extreme-white piebald gene (for which they are homozygous). The dominant ticking gene (T) Apunches@ through the white to show the black or liver. Dalmatians with blue eyes (strong piebald expression) are statistically more likely to be deaf. Dalmatians with a patch (weak piebald expression) are statistically less likely to be deaf. The same association between blue eyes and deafness holds in other dog breeds11 and most white cat breeds. Efforts through breeding to reduce blue eyes in Norwegian Dalmatians reduced the prevalence of deafness.19 Blue eyes are allowed in the Dalmatian breed standard of the United States, but not in Europe; deafness prevalence rates in Europe are approximately 20% compared to the US rate of 30%.
Deafness Prevalence
In dogs, congenital deafness has been reported in over 80 breeds (Table I). This deafness is not necessarily hereditary in all listed breeds, but must be assumed to be so in most cases. Deafness prevalence is known for several studied breeds, but is unknown for most breeds.11 For the few studied breeds, the prevalence rates are as follows:
|
Breed |
(N) |
Unilaterally deaf (%) |
Bilaterally deaf (%) |
Total deaf (%) |
|
Dalmatian |
(5,333) |
21.9 |
8.0 |
29.9 |
|
Bull terrier
white
colored |
(665)
(346)
(311) |
9.9
18.0
1.3 |
1.1
2.0
0.0 |
11.0
19.9
1.3 |
|
English setter |
(3,656) |
6.5 |
1.4 |
7.9 |
|
English cocker spaniel |
(1,136) |
5.9 |
1.1 |
6.9 |
|
Australian cattle dog |
(296) |
12.2 |
2.4 |
14.5 |
The Dalmatian is sw, the English setter is usually sp, the English cocker spaniel (based on the American cocker spaniel) is usually sp, and the white bull terrier is sw while the colored bull terrier is si;13 the allele in the cattle dog is not known.
It is not necessarily the case that prevalence rates are lower in breeds other than these. Rather, it is often the case that breeds with deafness problems have not yet adequately addressed the problem. Few similar deafness studies have been reported for other breeds, except for the Norwegian dunkerhound, in which unilateral or bilateral deafness has been reported to occur in 75% of all white animals,20 and the dappled (merle) dachshund, where 18.2% were reported to be bilaterally deaf and 36.4% were unilaterally deaf.21 In both breeds the prevalence of deafness in colored or non-dappled dogs was not documented but is low.
Deafness prevalence in white cats has been studied by several investigators (summarized in Delack6), but the best data come from the breeding studies of Mair7 and Bergsma and Brown16 who examined crosses between white and non-white parents and between hearing and deaf parents. Prevalence rates among white kittens for deafness in one or both ears were 51.5% (N=66) and 42.6% (N=162) respectively. When kittens were homozygous for white (WW) the rates were 96.0% and 52.0% in the two studies; heterozygotes were 24.3% and 27.4%. In the Mair study at least one parent was always bilaterally deaf, while Bergsma and Brown included all possible hearing combinations in the parents, making it difficult to compare results between the studies. From these complex studies it is difficult to cite a single prevalence rate, but clearly the prevalence rates are high. The authors also found a clear relationship between blue eyes and deafness: deafness was 3-5 times more common in cats with two blue eyes than cats without blue eyes, and two times more common in cats with one blue eye. Pure cat breeds carrying the W gene, potentially susceptible to deafness, are listed in Table II.
Genetics of Deafness
The inheritance of the pigment genes described above does not provide a simple description of the inheritance of the deafness that is so often associated with white pigmentation. Hereditary deafness can potentially result from any of several mechanisms: autosomal dominant. autosomal recessive, X-linked, mitochondrial, or polygenic (or multifactorial). In most instances the mechanism is unknown. Incomplete penetrance, where not all aspects of a deafness syndrome are expressed in an affected individual, frequently complicates an understanding of the mode of inheritance. To complicate the topic even further, environmental factors may interact with genetic factors to trigger expression of a disease.
No known X-linked or mitochondrial deafness has been reported in dogs or cats. Although it has been a controversial subject,22 there appears to be no gender difference in deafness prevalence.11 Many literature citations report deafness in piebald-carrying breeds to be autosomal recessive, but this is incorrect: the associated pigment gene is recessive but the deafness is not. It is common to observe deaf offspring from two normal parents, ruling out an autosomal dominant mechanism. In a research breeding colony of deaf Dalmatians at LSU, bilaterally hearing puppies often resulted from breeding two bilaterally deaf parents, ruling out a simple autosomal recessive mechanism since the deaf parents should have been homozygous for the responsible gene. The result is unresolved possibilities of incomplete penetrance of a recessive gene, multiple genes, or modifier genes acting on a single recessive gene. As terminology has evolved in genetics, and a better understanding of the molecular bases for gene actions develops, it may turn out that incomplete penetrance, polygenes, and modifier genes prove to result from the same molecular mechanism. When working to identify the physical location of a gene on a chromosome, targets are referred to as loci (singular: locus) when an approximate location on a single chromosome has been found but the specific gene is not yet identified. Because a locus consists of many genes, several of which may be involved in inheritance of the disease, it is often difficult to unequivocally explain the mode of inheritance until a single gene is identified and sequenced. A recent study with Dalmatians23 suggests that deafness is inherited as a single locus, but one that does not follow Mendelian genetics (see below). This finding appears to be supported by other studies.24,25 Nevertheless, except for the Doberman, the mechanism(s) of inheritance of deafness in dogs and cats is not yet known. It has been stated that white in the cat is a simple dominant gene with pleiotropic effects that include blue eyes and degeneration of the inner ear;6 it might also be said that the piebald alleles of S in dogs are simple recessive genes with pleiotropic effects that include blue eyes and degeneration of the inner ear (and likewise for merle), but neither statement provides a means to describe the inheritance of deafness.
The classic genetics of coat color in dogs is only now benefiting from molecular genetic studies, advancing toward a knowledge of which chromosome-based genes are responsible for the phenotype-based coat color genes S and M; studies in cats do not yet appear to be in progress. To date, several canine chromosomal genes have been excluded for S and M26,27 but no promising candidates have been identified. By far, the great majority of the breeds affected by hereditary deafness are carriers of recessive alleles of the phenotype S gene, so identifying its location will be important.
Deafness in Dobermans, which do not carry the merle or piebald genes, results from direct loss of cochlear hair cells without any effects on the stria vascularis B primary sensorineural deafness.28 Vestibular system signs, including head tilt and circling, are seen, and the deafness has been reported to be transmitted by a simple autosomal recessive mechanism. A similar pathology has been described for the Shropshire Terrier.
Until a gene mutation is identified for pigment-associated deafness, and a DNA-based diagnostic test developed, breeders must base breeding decisions on the hearing background of potential mates if deafness prevalence is to be reduced. Since unilateral deafness is genetically the same as bilateral deafness (but just not completely expressed), unilaterally deaf animals should not be bred B and of course bilaterally deaf animals should not be bred. Even when both parents are normal, it is unwise to repeat a breeding that produced deaf animals, although in breeds with a high prevalence rates this may be difficult. Breeding to animals with a history of producing deaf should be avoided. Breeding to blue-eyed animals should be avoided in most breeds. If deafness has been identified as a problem in a breed, the hearing status of the animals to be bred should always be known in advance. Finally, breeding decisions should always take into consideration the overall good of the breed.
Current Research
Significant effort is presently being invested in canine deafness studies in the US and elsewhere.
To initiate identification of a chromosomal locus that co-segregates with deafness in the Dalmatian, we have established an extended pedigree of 266 related Dalmatians, of which 199 have known hearing status (KE Murphy, Texas A&M University and GM Strain). Through use of a statistical technique known as complex segregation analysis we have shown from this kindred that deafness is best modeled as resulting from a single major locus with an affect on deafness, but that this locus does not appear to follow Mendelian inheritance.23 Thus, deafness results from one gene (or a locus of closely located genes that segregate together), but the inheritance does not appear to be simple autosomal recessive, which explains the difficulty of tracking deafness in pedigrees of affected animals. DNA has been isolated from 117 of these related Dalmatians for studies to isolate the genetic cause of deafness.
One molecular approach in identifying the cause of deafness is by examination of the canine equivalent of genes identified as being responsible for deafness in humans or mice (candidate gene approach). Not knowing the mechanism of inheritance of deafness has made these molecular genetic studies difficult. However, it has now been shown that several of the most promising genes are not at fault in the dog: pax3, mitf, endrb, c-kit, and kitlg.26,27,29,30 Literally hundreds of loci that cause deafness have been identified in humans and mice,31 and there is no guarantee that the gene or genes responsible for pigment-associated deafness in dogs have already been identified in the human or mouse. As a consequence, we have adopted the approach of performing a whole-genome screen of DNA from this established Dalmatian pedigree. Linkage analysis is being performed on our assembled Dalmatian DNA using 172 multiplexed microsatellite markers that span the canine genome at < 10 cM spacing.32,33 Similar pedigrees of DNA are being established from English setters and Jack Russell terriers, since any causative gene mutation identified in the Dalmatian must be examined for also being responsible for deafness in other affected breeds. Multiple different mutations in a single gene can produce the same disease, as has been shown for several human deafness syndromes, and mutations in several different genes can produce diseases that cannot be distinguished clinically.
In addition to the above studies, studies are currently underway at several other institutions. Hereditary deafness in Anervous@ pointers is being studied at the University of Pennsylvania, several candidate genes are being studied at Michigan State University, and candidate gene and whole genome screen studies of deafness are being pursued at several locations in Europe. No similar studies are underway in cats to the author's knowledge.
Table I. Breeds with reported congenital sensorineural deafness (from personal observations by the author, communications from breeders, and as summarized in Strain9).
|
Akita
American bulldog
American-Canadian shepherd
American Eskimo
American Staffordshire terrier
Australian cattle dog
Australian shepherd
Beagle
Bichon Frise
Border collie
Borzoi
Boston terrier
Boxer
Bulldog
Bull terrier
Cardigan Welsh Corgi
Catahoula leopard dog
Cavalier King Charles spaniel
Chihuahua
Chinese crested
Chow chow
Cocker spaniel
Collie
Coton de Tulear
Dalmatian
Dappled dachshund
Doberman pinscher |
Dogo Argentino
English bulldog
English cocker spaniel
English setter
Foxhound
Fox terrier
French bulldog
German shepherd
Great Dane
Great Pyrenees
Greyhound
Havanese
Ibizan hound
Italian greyhound
Jack Russell terrier
Kuvasz
Labrador retriever
Löwchen
Maltese
Miniature pinscher
Miniature poodle
Mongrel
Norwegian dunkerhound
Nova Scotia duck tolling retriever
Old English sheepdog
Papillon
Perro de Carea Leonés |
Pit bull terrier
Pointer
Presa Canario
Puli
Rhodesian ridgeback
Rat terrier
Rottweiler
Saint Bernard
Samoyed
Schnauzer
Scottish terrier
Sealyham Terrier
Shetland sheepdog
Shih Tzû
Shropshire terrier
Siberian husky
Soft coated Wheaten terrier
Springer spaniel
Sussex spaniel
Tibetan spaniel
Tibetan terrier
Toy fox terrier
Toy poodle
Walker American foxhound
West Highland white terrier
Whippet
Yorkshire terrier |
Table II. Cat breeds with the dominant white gene (W).34
|
White |
White Cornish rex |
|
White Scottish fold |
White Devon rex |
|
European white |
White British shorthair |
|
White Turkish angora |
White manx |
|
Foreign white |
White exotic shorthair |
|
White American wirehair |
White Persian |
|
White American shorthair |
White oriental shorthair |
References
1. Steel KP, Barkway C. 1989. Another role for melanocytes: Their importance for normal stria vascularis development in the mammalian inner ear. Development 107:453-63.
2. Strain GM, Tedford BL, Jackson RM. 1991. Postnatal development of the brainstem auditory-evoked potential in dogs. American Journal of Veterinary Research 52:410-5.
3. Strain GM, Green KD, Twedt AC et al. 1993. Brain stem auditory evoked potentials from bone stimulation in dogs. American Journal of Veterinary Research 54:1817-21.
4. Strain GM, Tedford BL, Littlefield-Chabaud MA, et al. 1998. Air- and bone-conducted brainstem auditory evoked potentials and flash visual evoked potentials in cats. American Journal of Veterinary Research 59:135-137.
5. Strain GM. Deafness in Dogs and Cats. http://www.lsu.edu/deafness/deaf.htm.
6. Delack JB. 1984. Hereditary deafness in the white cat. Compendium on Continuing Education for the Practicing Veterinarian 6:609-619.
7. Mair IWS. 1973. Hereditary deafness in the white cat. Acta Otolaryngologica Suppl 314:1-48.
8. Mair IWS. 1976. Hereditary deafness in the Dalmatian dog. Archives of Otorhinolaryngology 212:1-14.
9. Strain GM. 1996. Aetiology, prevalence, and diagnosis of deafness in dogs and cats. British Veterinary Journal 152:17-36.
10. Strain GM. 1999. Congenital deafness and its recognition. Veterinary Clinics of North America: Small Animal Practitioner 29:895-907.
11. Strain GM. 2003. Deafness prevalence and pigmentation and gender associations in dog breeds at risk. The Veterinary Journal (in press).
12. Strain GM, Kearney MT, Gignac IJ, et al. 1992. Brainstem auditory evoked potential assessment of congenital deafness in Dalmatians: associations with phenotypic markers. Journal of Veterinary Internal Medicine 6:175-82.
13. Little CC. 1957. The Inheritance Of Coat Color in Dogs. Howell Book House: New York. 194 pp.
14. Sponenberg DP, Rothschild MF 2001. Genetics of coat colour and hair texture. In The Genetics of the Dog, eds. A. Ruvinsky, J. Sampson, pp. 61-85. Wallingford, Oxon, UK: CABI Publishing.
15. Sponenberg DP. 1985. Inheritance of the harlequin colour in Great Dane dogs. Journal of Heredity 76:224-5.
16. Bergsma DR, Brown KS. 1971. White fur, blue eyes, and deafness in the domestic cat. Journal of Heredity 62:171-185.
17. Searle AG. 1968. Comparative Genetics of Coat Colour In Mammals. Logos Press/Academic Press: London. 310 pp.
18. Pedersen APG. 1991. Feline Husbandry. American Veterinary Publications: Goleta, CA.
19. Greibrokk T. 1994. Hereditary deafness in the Dalmatian: relationship to eye and coat color. Journal of the AmericanAnimalHospital Association 30:170-6.
20. Foss I. 1981. Development of hearing and vision, and morphological examination of the inner ear in hereditarily deaf white Norwegian dunkerhounds and normal dogs (black and dappled Norwegian dunkerhounds). M.Sc. Thesis, Cornell University, Ithaca, NY, 133 pp.
21. Reetz I, Stecker M, Wegner W. 1977. Audiometrische Befunde in einer Merlezucht. Deutsche Tierärztliche Wochenschrift 84:273-7.
22. Wood JLN, Lakhani KH. 1997. Prevalence and prevention of deafness in the Dalmatian B Assessing the effect of parental hearing status and gender using ordinary logistic and generalized random litter effect models. Veterinary Journal 154:121-33.
23. Cargill EJ, Famula TR, Strain GM, et al. 2003. Heritability and segregation analysis of deafness in US Dalmatians. Genetics (submitted).
24. Famula TR, Oberbauer AM, Sousa CA. 2000. Complex segregation analysis of deafness in Dalmatians. American Journal of Veterinary Research 61:550-3.
25. Muhle AC, Jaggy A, Stricker C, et al. 2002. Further contributions to the genetic aspect of congenital sensorineural deafness in Dalmatians. Veterinary Journal 163:311-8.
26. Metallinos D, Rine J. 2000. Exclusion of EDNRB and KIT as the basis for white spotting in Border Collies. Genome Biology 1:research0004.1-research0004.4.
27. Schmutz SM, Berryere TG, Sharp CA. 2003. KITLG maps to canine chromosome 15 and is excluded as a candidate gene for merle in dogs. Animal Genetics 34:75-76.
28. Wilkes MK, Palmer AC. 1992. Congenital deafness and vestibular deficit in the doberman. Journal of Small Animal Practice 33:218-224.
29. Brenig B, Pfeiffer I, Jaggy A, et al. 2003. Analysis of the 5' region of the canine PAX3 gene and exclusion as a candidate for Dalmatian deafness. Animal Genetics 34:47-50.
30. Zemke D, Cao Y, Yuzbasiyan-Gurkan V. 1999. Hereditary hearing loss in dogs: models for sensorineural deafness. Hereditary Deafness Newslet. 16:34 (www.ihr.mrc.ac.uk/ hereditary/newsletters/ index.shtml).
31. Van Camp G, Smith RJH. Hereditary Hearing Loss Homepage. http://dnalab-www.uia.ac.be/dnalab/hhh/.
32. Cargill EJ, Clark LA, Steiner JM et al. 2002. Multiplexing of canine microsatellite markers for whole-genome screens. Genomics 80:250-3.
33. Richman M, Mellersh CS, André C, et al. 2001. Characterization of a minimal screening set of 172 microsatellite markers for genome-wide screens of the canine genome. Journal of Biochemical and Biophysical Methods 47:137-49.
34. Gebhardt RH, Pond G, Raleigh I. 1979. A Standard Guide to Cat Breeds. McGraw-Hill Book Co: New York.