Pacific Identifications, Inc.
Victoria, BC, Canada
Introduction
If we accept the premise that dogs were brought to the Americas from Eurasia by their human companions (or followed along uninvited)-rather than domesticated here from local wolves--then we might expect North American (NA'an) archaeological sites to yield remains of the oldest dogs as well as those with the greatest diversity in form. In this broad review of the history of dogs in North America (NA) before European contact, I address these assumptions and discuss their broader implications. I include a summary of the dates for the oldest dog remains recovered so far in the Americas and Eurasia. The evidence for the development of discrete dog varieties in North America includes an in-depth account of the little-known Salish Wool dog of the central Northwest Coast (NWC), which may be the only true breed (as defined by modern standards) developed prehistorically in the Americas.
when did dogs come to north america?
Dogs are the very first domestic animals to appear in the archaeological record. We have evidence from at least 14,000 years ago of fully domesticated dogs in Eurasia (Figure 1)(1-16). These early dogs share several features, including a much shortened facial region, crowded teeth, and smaller overall size compared to contemporaneous local wolves(4).
Figure 1. Dates and locations of early dog remains worldwide.
|
Continent |
Country |
Locale |
Material |
Dates (uncorrected) |
|
Europe |
Germany |
Kniegrotte Cave |
skull frag |
12,280 ± 90,
13,585 ± 165 BP,
on strata (1) |
|
|
Germany |
Oberkassel |
mandible |
ca. 14,000 BP,
on strata (2) |
|
|
Germany |
Bedburg-Köningshoven |
skull (adult) |
ca. 10,000 BP,
on strata (3) |
|
|
England |
Star Carr
Seamer Carr |
skull frag
(juvenile)
verts (juv.) |
9,559 ± 210, on strata,
9,490 ± 350, on strata
9,940 ± 110 BP,
on bone (4) |
|
|
France |
St. Thibaud
Rockshelter (Alps) |
skull +
femur, vert. |
10,050 ± 100 BP,
on bone (5) |
|
W. Asia |
Israel |
Ein Mallaha |
full skeleton
(burial; juv.) |
ca. 12,000 BP,
on strata (6) |
|
|
Israel |
Hayonim |
full skeletons
(2; burials) |
ca. 12,500 BP,
on strata (7) |
|
|
Russia (West, near Kiev) |
Eliseevichi I (53° N, 33°E) |
skulls (adult) (2) |
13,900 ± 55 BP,
on bone (8,9) |
|
|
Russia (West, Altai region) |
Razboinichiya Cave
(51°N, 85°E) |
skull |
14,850 ± 70 BP,
on strata (10, 11) |
|
E. Asia |
China (North) |
Jiahu (Wuyang, Henan); |
full skeletons
(11; burials) |
ca. 9,000-7,800 BP,
on strata* |
|
|
China (North) (preliminary) |
Nanzhunangtou,
(Xushui, Hebei) |
mandible |
ca. 10,000 BP,
on strata1 |
|
|
Japan |
Natsushima Shell
Mound (Kanagawa) |
not specified |
ca. 9,300 BP,
on strata (12) |
|
|
Japan |
Kamikuroiwa
Rockshelter (Ehima) |
full skeleton
(burial) |
ca. 8,500-8,000 BP,
on strata (12, 13) |
|
|
Russia (Far East, on Kamchatka) |
Ushki-I |
full skeleton (burial) |
10,360 ± 45,
10,860 ± 40 BP, on strata (14) |
|
N. America |
USA (Utah) |
Danger Cave |
mandibles & skull frags |
ca. 9,000-10,000 BP,
on strata (15) |
|
|
USA (Illinois) |
Koster |
full skeletons
(3; burials) |
8,130 ± 90,
8,480 ± 110 BP, on strata (16) |
*Dr. Yuan Jing, personal communication, 2004. Chinese Academy of Social Sciences, Institute of Archaeology (yuanj0091@sina.com).
Despite what all of us have been taught about domestication, scientists understand very little about the precise biological and/or anthropogenic mechanisms responsible for transforming wild animals into domestic ones(17-22). This has important implications for our understanding of breed development because many assumptions have been made about the nature of the early dog/human relationship that are probably incorrect, as has been shown for long-held assumptions regarding domestication itself(20,42). Certainly, there is no evidence to suggest that deliberate human actions precipitated domestication in most animals and the present consensus of opinion is that domestication was initiated by the animals themselves(20,23-25). This conclusion is supported by evidence from recent genetic studies that domestication of wild ancestral species has occurred more than once, in different parts of the world, for virtually all domestic animals(26-40). For example, dogs appear to have been generated on at least three separate occasions (perhaps more) from geographically distinct ancestral populations of wolf (i.e., different subspecies). A similar pattern is seen in domestic pigs, cattle, sheep, goats, horses and water buffalo; all but the chicken show evidence for two or more discrete domestication events in different geographic locations.
Such multiple domestication events from geographically distinct subspecies of wolf could perhaps account for some of the variation we see amongst early prehistoric dogs. It is important to separate such initial variation from variation generated much, which is the result of local adaptation and/or breed creation. Although no comprehensive study of dogs from different geographic regions has yet been done, an examination of published data indicates that early dogs worldwide were remarkably similar in every way except size (42). Early dogs from northern Europe and Russia were the largest (although no larger than a modern dingo), while most from southwest Asia, China and NA were Dalmation-sized. All early dogs from Japan and one from central Europe (5) were somewhat smaller (about the size of a modern Finnish spitz or Keeshound). All were robust and well-proportioned but similar in general conformation: in all cases, slight differences in size are virtually all that distinguish dogs for thousands of years regardless of where they lived (1-16, 42-44).
Early dogs in NA were considerably smaller than any NA'an subspecies of wolf and it has been suggested, based in part on a distinct shared feature of the mandible, that a smaller wolf subspecies from Eastern Asia was ancestral to NA'an dogs(45-47). NA'an dogs must have accompanied human migrants on one or several of their journeys from Asia, an assumption now supported by DNA analysis of ancient dogs from the Americas(26-29). However, it is probable that some NA'an dogs originated from different Asian regions and entered NA at different times.
Breed development in North America
In Eurasia, archaeological evidence for the deliberate differentiation of dogs into discrete breeds appears relatively late. Large dog types appear first, during the Iron Age, about 4,000 years ago (48). Controlled breeding and deliberate selection of heritable traits are all that is needed to produce a true domestic breed within a few generations, although it has been suggested that a written language is also needed to keep breeding records(42).
The Romans appear to have used the knowledge gained from the experience of producing large dogs via controlled breeding to develop other distinct dog types. Small dogs, even lap dogs, appear about 3,000 years ago and spread quite rapidly throughout the Roman Empire (48-51). Although such breed development probably occurred in Egypt and China about this time or even earlier as well, archaeological records pertinent to addressing these questions is either rare or unavailable to Western researchers(42).
In NA, the picture is quite different. While dogs were clearly abundant, none of these societies had a written language. Early native dogs of NA (ca. 10,000-4,000 BP) show remarkable similarity in size and general conformation: all found so far resemble the three oldest complete dog burials from Illinois (all large, about the size of a modern Dalmatian) (16, 52-53). Marked differences in size are not apparent until after about 4,000BP. More than 60 years ago, William Haag documented some of this variation in a comprehensive osteometric analysis of archaeological dog remains(54); more samples have been analyzed and reported since then(55-59).
Amongst dog samples recovered from the last 4,000 years, some regions show a wide range of variation in size while others do not (small dogs have been defined as those with a total skull length of 108-165 mm, accompanied by a humerus total length below 140 mm and femur total length below 160 mm; large dogs are defined as those with a total skull length of 165-196 mm, with humerus length above 140 mm and femur length above 160 mm)(55). For example, archaeological sites in Alaska and Greenland yield predominantly large dogs (although no larger than a modern dingo), while sites in the Kentucky/Alabama region generate mostly small ones(16, 59-60). While different sizes of dogs existed side by side in many regions (such as the US southwest and at the Jaguar Cave site in Idaho)(47), there is little evidence to suggest that populations of small dogs were the result of deliberate selective breeding. In only one of these regions is there associated evidence to suggest that conscious selection for distinct physical attributes was accompanied by controlled breeding--in other word, the necessary elements required to produce a truly distinct breed of dog as defined by modern standards.
Northwest Coast Wool Dog--a true native breed
Could there have been true husbandry for specific breeds in NA before there were written records to keep track of individual relationships and lineages? I suggest that on the central NWC of North America, a good example exists of such prehistoric breeding, summarised below from my own research on archaeological remains and several other comprehensive accounts(55). In contrast to other NA'an archaeological samples where size variation in skeletal remains has been reported, or where ethnographic reports suggest different types of dogs existed(54,56,62-64), on the NW Coast there is strong ethnographic evidence for actual dog husbandry. These accounts describe two types of dogs deliberately maintained as separate populations, based on distinct heritable morphological traits, with an explicitly stated economic rationale strong enough to overcome the practical difficulties of dog husbandry without benefit of non-chewable restraints and fencing materials.
The early historic and ethnographic accounts report the presence of two types of native dogs kept by aboriginal peoples on the central NWC of NA: a medium-sized, coyote-like animal (the "Village dog") and a smaller, long-haired dog (the "Wool dog"). The Wool dog had a very limited distribution within a small region of southern British Columbia (including southeastern Vancouver Island, the Gulf Islands and the lower Fraser River) and northern Washington State (including Puget Sound and the Olympic Peninsula) (Figure 2).
| Figure 2. | 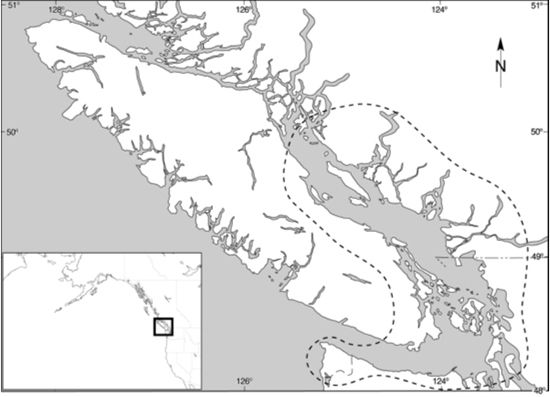
Map of the central NWC of NA (insert), with dashed line showing the limited region (southern Vancouver Island/lower Fraser River/Georgia Straight/Puget Sound/Olympic Peninsula) where archaeological remains of putative wool dogs have been found or were reported by early explorers (courtesy I. McKechnie). |
|
| |
The Wool dog apparently existed in rather substantial numbers in the late 1700's and early 1800's when Europeans descended on this region. The Wool dog was observed by these early explorers to serve an important purpose in the native economy: their wool was spun (often combined with other ingredients) and woven on impressive standing looms into ceremonial blankets (66). These handsome woven blankets provided the incentive for breeding Wool dogs because without dog's wool, the blankets could not be produced in any quantity. Native NA'ans of the NWC had no other source of wool except the wild mountain goat, which lived high in the northern coastal and interior mountains. Mountain goat wool was a rare commodity that was used extensively only by tribes who had easy access to the mountains; other groups paid a high price in trade for mountain goat wool.
The problem with keeping Wool dogs as a "sheep surrogate" was that another variety of dog also existed. The common "Village dog" was similar in size and form to other pariah-type dogs world-wide (e.g., Allen's "Plains Indian dog," [62]), somewhat smaller than a modern dingo. Village dogs were common in many communities and although a few may have been trained to assist in hunting, most were not owned or cared for in any way. The Village dog was not only taller than the Wool dog, it also had short hair (see Figure 3). We now know that long hair is recessive to short hair and that first-generation hybrids between these two extremes generally possess hair of intermediate length. This suggests that even without a knowledge of genetics and inheritance, the effect of interbreeding the two types would have been readily apparent, since hybrid offspring would resemble the shorthaired village dog more than the longhaired wool dog in coat type. Such hybrids between the two types would be immediately and continuously identifiable throughout the dog's life, even without written records. Without the desired long woolly coat it is doubtful that hybrid offspring between the two types would have been used for future deliberate breeding.
| Figure 3. | 
Artistic reconstruction of NWC Village dog (left, shoulder height ca. 52 cm) and Wool dog (right, shoulder height ca. 44 cm), scaled to show relative size of each; drawings by C.J. Pye (55, 67). |
|
| |
The recessive inheritance pattern of the desired trait must have been the key factor that made selective breeding possible, making it obvious that keeping individuals with long hair reproductively isolated from others was necessary for the continued production of longhaired dogs. In order to maintain reproductive isolation, wool dogs were said to be left on a small island, with dried food buried for the dogs to find, when they could not be closely watched by their owners.
Keeping dog numbers under control would have been especially important where two breeds needed to be kept apart. One method of population control for dogs that has been described for indigenous people from other areas is to limit the number of breeding females--that is, to allow only as many females to reach sexual maturity as is needed to maintain the population. This is said to effectively reduce the particularly vicious fighting that occurs between males when females are reproductively receptive ("in heat"). If this was true on the NWC, we would expect to find relatively large numbers of adult, breeding age females of the Wool dog type (as an indication that females of that type were valued and were being deliberately kept for breeding). As the population of larger Village dogs was not managed in the same way, the adult portion of that population should have been almost exclusively male. In fact, this is precisely what has been found, at least for the last 1400 years (almost equal numbers of adult male and female putative Wool dog remains vs. mostly male putative Village dog remains). See Figure 4 for a comparison of both types to other prehistoric dog samples.
Brightly striped Hudson's Bay blankets were common trade goods during the early period of European contact and the attractive goods encouraged most aboriginal weavers to abandon the making of Wool dog blankets. As the economic value of dog's wool declined, Wool dogs were allowed to interbreed freely with both native Village dogs and with any European dogs brought into communities by early settlers. By 1858, Wool dogs were considered extinct as a distinct breed. Although commercial sheep's wool soon became available and weaving on the traditional native looms resumed, by that time the distinctive Wool dog was lost forever.
Figure 4. Selected measurements of NWC dogs vs. other prehistoric samples (30).
|
Dimensions
(mm) |
Wool Dog
(55) |
Village Dog
(55) |
Kentucky
(54) |
Japanese
Jomon (68) |
Peruvian
(68) |
Preboreal
France (5) |
|
|
(NWC small) |
(NWC large) |
|
|
|
10,000 BP |
|
|
Mean |
Mean |
Mean |
Mean |
Mean |
|
|
Skull |
n (6-10) |
n (7-9) |
n (27-50) |
n (4-8) |
n (21-25) |
n (1) |
|
Total length |
162.0 |
188.6 |
153.9 |
163.0 |
159.8 |
149.0 |
|
Condylobasal
length |
154.6 |
172.1 |
145.6 |
|
|
140.5 |
|
|
(NWC small) |
(NWC large) |
Kentucky |
Jomon |
|
Preboreal |
|
Longbone
Elements |
n (10-20) |
n (10-21) |
n (43-101) |
n (5-9) |
|
n (1) |
|
Femur GL |
154.3 |
175.3 |
129.3 |
134.8 |
|
130.4 |
|
Tibia GL |
150.0 |
165.7 |
127.5 |
141.8 |
|
|
|
Humerus GL |
143.5 |
161.3 |
118.0 |
131.3 |
|
|
|
Radius GL |
136.0 |
149.9 |
115.8 |
125.7 |
|
|
|
Ulna GL |
157.5 |
180.8 |
143.3 |
148.8 |
|
|
References
1. Musil, R. Domestication of wolves in central European Madalenian sites. In: Dogs Through Time: In Archaeological Perspective, S. J. Crockford (ed). Archaeopress, B.A.R. S889 Oxford, 2000: 21-28.
2. Nobis, G. 1979, cited in (4).
3. Streeet, M. 1989, cited in (4).
4. Clutton-Brock, J. Origins of the dog: domestication and early history. In: The Domestic Dog: Its Evolution, Behaviour and Interactions with People, J. Serpell (ed.). Cambridge University Press Cambridge, 1995: 8-20.
5. Chaix, L. A preboreal dog from the northern Alps (Savoie, France). In: Dogs Through Time: An Archaeological Perspective, S. J. Crockford (ed). Archaeopress, B.A.R. S889 Oxford, 2000: 49-59.
6. Davis, S. J. M. and F. R. Valla. Evidence for domestication of the dog 12,000 years ago in the Natufian of Israel. Nature 1978, 276:608-610.
7. Tchernov, E. and F. F. Valla. Two new dogs, and other Natufian dogs, from the Southern Levant. Journal of Archaeological Science 1997, 24:65-95.
8. Sablin, M.V. and G.A. Khlopachev. The earliest ice age dogs: evidence from Eliseevichi I. Current Anthropology 2002, 43(5):795-799.
9. Sablin, M. V. and G. A. Khlopachev. Die altesten hunde aus Eliseevici I (Russland). Archaologisches Korrespondenzblatt 33. Romish-Germanischen Zentralmuseum, Mainz, Germany, 2003.
10. Ovodov, N. D. The ancient dogs of Siberia. Abstracts of the 8th Congress of the International Council for Archaeozoology(August 23-29), Victoria, British Columbia, Canada, 1998.
11. Turner II, C. G. Teeth, needles, dogs, and Siberia: Bioarchaeological evidence for the colonization of the New World. In: The First Americans: Pleistocene Colonization of the New World, N. G. Jablonski (ed.). Memoirs of the California Academy of Sciences 27, 2002: 123-158.
12. Shigehara, N. and H. Hongo. Ancient remains of Jomon dogs from Neolithic sites in Japan. In: Dogs Through Time: An Archaeological Perspective, S. J. Crockford (ed). Archaeopress, B.A.R. S889 Oxford, 2000: 61-67.
13. Shigehara, N. Morphological changes in Japanese ancient dogs. Archaeozoologica 1994, 6: 79-94.
14. Dikov, N. N. 1979. cited in Novikov, A. V. Dogs in the system of views and ritual practice of ancient populations of west Siberian forest-steppe and south-taiga zones. Archaeology, Ethnology and Anthropology of Eurasia 2001, 1(5): 72-83.
15. Grayson, D. K. Danger Cave, Last Supper Cave, and Hanging Rock Shelter: The faunas. Anthropological Papers of the American Museum of Natural History 66(1), 1988.
16. Morey, D. F. and M. D. Wiant. Early Holocene domestic dog burials from the North American midwest. Current Anthropology 1992, 33: 224-229.
17. Diamond, J. Guns, Germs and Steel: The Fates of Human Societies. W.W. Norton & Co., New York, 1999.
18. Reed, C. A. The beginnings of animal domestication. In: Evolution of Domesticated Animals, I.L. Mason (ed.). Longman Co., London 1984: 1-6.
19. Gould, S. J. A dog's life in Galton's polyhedron. Eight Little Piggies: Reflections in Natural History, W. W. Norton and Co., New York. 1994: 382-395.
20. Crockford, S. J. Animal domestication and vertebrate speciation: a paradigm for the origin of species. Doctoral dissertation, University of Victoria, British Columbia, 2004.
21. Coppinger, R. and R. Schneider. Evolution of working dogs. In: The Domestic Dog: Its Evolution, Behaviour and Interactions with People, J. Serpell (ed.). Cambridge University Press Cambridge, 1995: 21-47.
22. McKinney, M. L. The juvenilzed ape myth-our "overdeveloped" brain. BioScience 1998, 48: 109-116.
23. Tchernov, E. and L. K. Horwitz. Body size diminution under domestication: unconscious selection in primeval domesticates. Journal of Anthropological Archaeology 1991, 10: 54-75.
24. Morey, D. F. The early evolution of the domestic dog. American Scientist 1994 (July/Aug): 336-347.
25. Coppinger, R. and L. Coppinger. Dogs: A New Understanding of Canine Origins, Behavior and Evolution. University of Chicago Press, Chicago, 2001.
26. Koop B. F., M. Burbidge, A. Byun, U. Rink and S. J. Crockford. Ancient DNA evidence of a separate origin for North American indigenous dogs. In: Dogs Through Time: An Archaeological Perspective, S. J. Crockford (ed). Archaeopress, B.A.R. S889 Oxford, 2000: 271-286.
27. Vilá, C., P. Savolainen, J. Maldonado, I. Amorim, J. E. Rice, R. L. Honeycutt, K. A. Crandall, J. Lundeberg and R. K. Wayne. Multiple and ancient origins of the domestic dog. Science 1997, 276: 1687-1689.
28. Morell, V. The origin of dogs: running with the wolves. Science 1997, 276: 1647-1648.
29. Leonard, J. A., R. K. Wayne, J. Wheeler, R. Valadez, S. Guillén, and C. Vilà. Ancient DNA evidence for Old World origin of New World dogs. Science 2002, 298: 1613-1616.
30. .MacHugh, D. E. and D. G. Bradley. Livestock genetic origins: Goats buck the trend. Proceedings of the National Academy of Sciences USA 2001, 98: 5382-5384.
31. Hiendleder, S., B. Kaupe, R. Wassmuth and A. Janke. Molecular analysis of wild and domestic sheep questions current nomenclature and provides evidence for domestication from two different subspecies. Proceedings of the Royal Society of London B 2002, 269: 893-904.
32. Luikart, G., L. Gielly, L. Excoffier, J.-D. Vigne, J. Bouvet, and P. Taberiet. Multiple maternal origins and weak phylogeographic structure in domestic goats. Proceedings of the National Academy of Sciences USA 2001, 98: 5927-5932.
33. Giuffra, E., J. M. H. Kijas, V. Amarger, O. Carlborg, J-T. Jeon and L. Andersson. The origin of the domestic pig: independent domestication and subsequent introgression. Genetics 2000, 154: 1785-1791.
34. Watanobe, T., N. Ishiguro, and M. Nakano. Phylogeography and population structure of the Japanese wild boar Sus scrofa leucomystax: mitochondrial DNA variation. Zoological Science 2003, 20: 1477-1489.
35. Watanobe, T., N. Ishiguro, M. Nakano, A. Matsui, H. Hongo, K. Yamazaki, and O. Takahashi. Prehistoric Sado Island populations of Sus scrofa distinguished from contemporary Japanese wild boar by ancient mitochondrial DNA. Zoological Science 2004, 21: 219-228.
36. Bradley, D. G., R. T. Loftus, P. Cunningham, and D. E. MacHugh. Genetics and domestic cattle origins. Evolutionary Anthropology 1998, 6: 79-86.
37. Mannen h., S. tsuji, R. T. Loftus and D. G. Bradley. Mitochondrial DNA variation and evolution of Japanese black cattle (Bos taurus). Genetics 1998, 150: 1169-1175.
38. Troy, C. S., D. E. MacHugh, J. F. Bailey, D. A. Magee, R. T. Loftus, P. Cunningham, A. T. Chamberlain, B. C. Sykes, and D. G. Bradley. Genetic evidence for Near-Eastern origins of European cattle. Nature 2001, 410: 1088-1091.
39. Jansen, T., P. Forster, M. A. Levine, H. Oelke, M. Hurles, C. Renfrew, J. Weber, and K. Olek. Mitochondrial DNA and the origins of the domestic horse. Proceedings of the National Academy of Sciences USA 2002, 99: 10905-10910.
40. Fumihito, A., T. Miyake, S-I. Sumi, M. Takada, S. Ohno, and N. Kondo. One subspecies of the red junglefowl (Gallus gallus gallus) suggices as the matriarchic ancestor of all domestic breeds. Proceedings of the National Academy of Sciences USA 1994, 91: 12505-12509.
41. Gentry, A., J. Clutton-Brock, and C. P. Groves. The naming of wild animal species and their domestic derivatives. Journal of Archaeological Science 2004, 31: 645-651.
42. Crockford, S. J. A commentary on dog evolution: regional variation, breed development and hybridization with wolves. In: Dogs Through Time: An Archaeological Perspective, S. J. Crockford (ed). Archaeopress, B.A.R. S889 Oxford, 2000: 295-312.
43. Dayan, T. and E. Galili. A preliminary look at some new domesticated dogs from submerged Neolithic sites off the Carmel coast. In: Dogs Through Time: An Archaeological Perspective, S. J. Crockford (ed). Archaeopress, B.A.R. S889 Oxford, 2000: 29-33.
44. Shigehara, N., Q. Guoqin, H. Komiya & Y. Jing. Morphological study of the ancient dogs from three Neolithic sites in China. International Journal of Osteoarchaeology 1998, 8: 11-22.
45. Weidensaul, S. Tracking America's first dogs. Smithsonian Magazine 1999, 29 (March): 44-57.
46. Olsen, S. J. and J. W. Olsen. The Chinese wolf, ancestor of New World dogs. Science 1977, 197: 533-535.
47. Olsen, S. J. Origins of the Domestic Dog: The Fossil Record. University of Arizona Press Tucson, 1985.
48. De Grossi Mazzorin, J. and A. Tagliacozzo. Morphological and osteological changes in the dog from the Neolithic to the Roman period in Italy. In: Dogs Through Time: An Archaeological Perspective, S. J. Crockford (ed). Archaeopress, B.A.R. S889 Oxford, 2000: 141-161.
49. Bartosiewicz, L. Metric variability in Roman period dogs in Pannonia Provence and the Barbaricum, Hungary. In: Dogs Through Time: An Archaeological Perspective, S. J. Crockford (ed). Archaeopress, B.A.R. S889 Oxford, 2000: 181-189.
50. Clark, K. M. Dogged persistence: the phenomenon of canine skeletal uniformity in British prehistory. In: Dogs Through Time: An Archaeological Perspective, S. J. Crockford (ed). Archaeopress, B.A.R. S889 Oxford, 2000: 163-169.
51. Cram, L. Varieties of dog in Roman Britain. In: Dogs Through Time: An Archaeological Perspective, S. J. Crockford (ed). Archaeopress, B.A.R. S889 Oxford, 2000: 171-180.
52. Cannon, A. Marine-based subsistence trends and the stable isotope analysis of dog bones from Namu, British Columbia. Journal of Archaeological Science 1999, 26: 399-407.
53. Yohe II, R.M. and M. G. Pavesic. Early Archaic dogs from the North American southeast. In: Dogs Through Time: An Archaeological Perspective, S. J. Crockford (ed). Archaeopress, B.A.R. S889 Oxford, 2000: 93-104.
54. Haag, W. G. An Osteometric Analysis of Some Aboriginal Dogs. University of Kentucky Reports in Anthropology 7, 1948.
55. Crockford, S.J. Osteometry of Makah and Coast Salish dogs. Archaeology Press 22, Simon Fraser University, Burnaby Canada, 1997.
56. Hadley, B.M. Preliminary results in determining dog types from prehistoric sites in the northeastern United States. In: Dogs Through Time: An Archaeological Perspective, S. J. Crockford (ed). Archaeopress, B.A.R. S889 Oxford, 2000: 205-215.
57. Morey, D. F. and K. Aaris-Sørensen. Paleoeskimo dogs of the Eastern Arctic. Arctic 2002, 55(1): 44-56.
58. Crockford, S., Letchford, B. and C. Moyer. Osteometric vs. genetic characterization of the Tahltan Bear Dog. In: Transitions in Zooarchaeology: New Methods and New Results. K. M. Stewart and F. L. Stewart (eds), Canadian Zooarchaeology Supplement #1, Canadian Museum of Nature Ottawa, 2003: 18-39.
59. Morey, D. F. Studies on Amerindian dogs: taxonomic analysis of canid crania from the northern Plains. Journal of Archaeological Science 1986, 13: 119-145.
60. Gollan, K. Prehistoric dingo. Doctoral dissertation, Australian National University, Canberra, 1982.
61. Clutton-Brock, J. Domesticated Animals From Early Times. London: British Museum (Natural History)/ Heinemann, 1981.
62. Schwartz, M. A History of Dogs in the Early Americas. Yale University Press New Haven, 1997.
63. Allen, G. M. Dogs of the American aborigines. Harvard University Museum of Comparative Zoology Bulletin 1920, 63: 431-517.
64. Walker, D.N. Preliminary bibliography on dogs and wolves, stressing their prehistoric and historic occurrence, hybridization, and domestication. In: Dogs Through Time: An Archaeological Perspective, S. J. Crockford (ed). Archaeopress, B.A.R. S889 Oxford, 2000: 313-343.
65. Wilcox, B. W. and C. Walkowicz. The Atlas of Dog Breeds of the World. T.F.H Publications Neptune City, 1989.
66. Amoss, P. T. Hair of the dog: unravelling pre-contact Coast Salish social stratification. In: American Indian Linguistics and Ethnography in Honor of Laurence C. Thompson. A. Mattina and T. Montler (eds.), University of Montana Missoula, 1993: 3-35.
67. Crockford, S. J. and C. J. Pye. Forensic reconstruction of prehistoric dogs from the Northwest Coast of North America. Canadian Journal of Archaeology 1997, 21: 149-153.
68. Clark, G. R. The kuri in prehistory: a skeletal analysis of the extinct Maori dog. MA thesis, University of Otago, New Zealand, 1995.